Regarding the explanations behind the diversity of exoplanets its also probably important to account for the innate biases in our assumptions and models for planet formation we also have to factor in planet evolution as well since there is pretty strong evidence that the conditions on Venus, Earth, Mars and even the orbital dynamics of Saturn have changed considerably since they initially formed, in fact all the planets as we know them are still dynamically changing thus was we see around older stars may not be representative of their formation conditions.
There was an interesting paper Anton Petrov brought to attention some years back with looked at the Kepler catalog and looked at the distribution of planets in size and compared that with their estimated age. While still obviously quite crude one standout result was that younger stars (those less than a billion years old) had larger diameter planets particularly an overabundance of "Mini Neptunes" whereas older stars (those more than a billion years old) show a higher abundance of "super Earths". The paper concluded that atmospheric escape is probably the best explanation for the difference.
This if verified by future work may also explain some of the extremely dense massive close in planets around old M dwarfs since if a gaseous world was evaporated its compressed rocky core might have become crystalized in such a form compensating for lower astrophysical metal abundances of their host stars.
From the perspective of geology however is where I come to support this kind of model most because as we have developed seismic tomography to probe the Earth's interior structure and began better characterizing magma chemical compositions and temperatures it has become apparent that there is significant thermochemical activity occurring deep inside our planet which interestingly has some surprisingly strong temporal cross matching to major ecological and climatic shifts in our planets history.
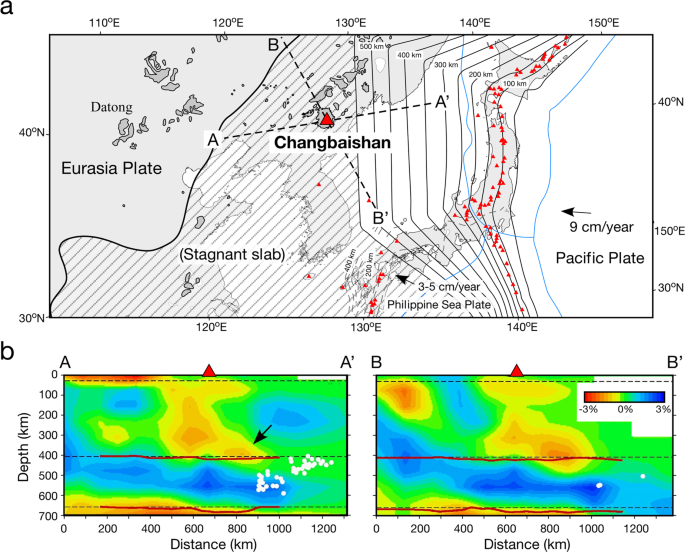
Magmatism at some intraplate volcanoes and large igneous provinces (LIPs) in continental areas may originate from hydrous mantle upwelling (i.e. a plume) from the mantle transition zone (MTZ) at 410–660 km depths in the Earth’s deep interior. However, the ultimate origin of the magmatism, i.e...

www.nature.com
In particular the temperature dependence of the phase transition from silicate rocks to hydrated silicates appears to be inversely proportional meaning that the reaction is inhibited in hot conditions probably in part because such processes are exothermic releasing heat thus the hotter Earth's mantle the less mineral hydrates can exist thus the more water will be able to exist in liquid form. This may help explain some curious observations related to the Early Earth namely the rate at which Earth's oceans formed, the lack of chemical isotopic signatures of atmospheric rock geochemistry prior to 3 billion years ago(Ga), and the peculiar observation based on geophysical evidence such as paleomagnetism fossils mineral types with time that the rate of motion and depths involved in plate tectonics on Earth have increased over time rather than decreased as would be expected were it purely fueled by thermal processes.
The early oceans can be explained if all that water was once in the oceans since given the sheer quantity it could have naturally allowed the water vapor in the aftermath of the Proto-earth Theia collision to have high enough temperatures and pressures to become supercritical fluids. This would mean there is no need for condensation type phase transitions since supercritical fluids don't undergo hard phase transitions able to naturally relax into a mix of liquid and gas phases in relatively equal proportions. In this case you could naturally have supercritical water oceans form on top of the still molten Earth before the crust even begins to solidify. This isn't far fetched as in the deep sea around hydrothermal vents supercritical water naturally exists on modern Earth so its existence on a hotter deeper ocean world is kind of trivial in this sense water would have behaved more like it does in Uranus and Neptune than the familiar surface water, plus the interface between a roiling molten magma ocean and a supercritical water ocean could be a place where exotic chemistry can potentially occur such as the reaction between iron and nickel rich volcanic glass(which is a solid which has frozen into the chemical structure of a liquid) which has been shown to readily catalyze the formation of RNA. could some more extreme chemical analog have seeded life on Earth?
In fact there is likely a deeper biological connection to this cycle as the 3 geophysical states of plate tectonics which we find in the rock record also correspond roughly to the Paleozoic + late Neoproterozic with "modern" plate tectonics and high levels of atmospheric oxygen, the Mesoproterozoic and Paleoproterozoic with a dynamic yet slower more shallow/less vigorous kind of tectonics and small percentages of oxygen in the atmosphere or shallow environments but otherwise completely anaerobic world often called the "boring billion" and then the old Archean eon and older which seem to have behaved even more unusually and for which there is evidence that while the continents physically existed to some degree they were likely fully submerged.
The following paper using modern analogs of microbial metabolisms explains an ecological reason for why these 3 different epochs of life each of which appears to have been divided by extreme and rapid transitions between a "snowball" and hot house state.
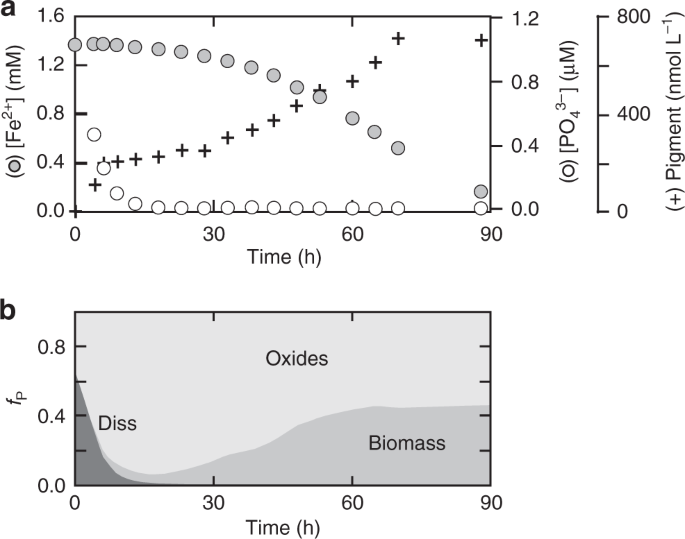
Competition dynamics between early Earth photosynthetic microorganisms are unclear. Here, the authors demonstrate that competition for light and nutrients between oxygenic phototrophs and Fe-based photosynthesizers in surface oceans provides a novel ecophysiological mechanism for the protracted...
www.nature.com
Effectively aerobic carbon fixation is ecologically limited by competition since it requires more energy to use water as a source of molecular hydrogen for carbon fixation. Thus in a water world where upwards passive diffusion is the primary source of mineral nutrients other methods of obtaining hydrogen for carbon fixing aerobic primary production is ecologically nonviable as nutrients will be exhausted by other anaerobic forms of photosynthesis which can use less energetic light before they can rise to the depths at which aerobic photosynthesis (which either requires at least 3 "red" photons or 1 "blue" to successfully split water into oxygen and molecular hydrogen) is possible.
This comes down to oxygen having the second highest electronegativity of all chemical elements that are not noble gasses and thus is the flip side of what makes aerobic respiration so efficient.
As a result aerobic carbon fixation requires too much energy for chemical processes or longer wavelengths of light (particularly infrared light) which are targeted by other kinds of anaerobic photosynthesis thus only in very shallow waters where sunlight can reach the bottom(or where some process saturates the water with abundant nutrients to such a degree that they are no longer a growth limiting factor) can aerobic photosynthesis become ecologically viable.
The aforementioned mantle hydration threshold happens to line up suspiciously well with the timing when such shallow water conditions would have been possible assuming all of Earth's water was initially in our oceans and has been subsequently sequestered away through crustal and mantle hydration processes. And that happens to line up within a few hundred million years or less of the Great Oxygenation Event(GOE).
Now as for why oxygen levels plateaued soon after the GOE, it turns out that while models have long assumed that anaerobic organisms can't stand oxygen that is only partly true since it has to do with oxygen saturation levels there turns out to be a lot of modern bacteria primarily in the soil and midwater depths which not only engage in anaerobic primary production/carbon fixation but use aerobic respiration primarily or even in some cases exclusively.
From what I have read these bacterial groups are actually extremely abundant at least in pelagic environments where they account for at least 20% to the total bacterial biomass in Earth's oceans. Its hard to know exactly how abundant they are because unfortunately like most microorganisms aside from laboratory favorites which have been studied to death microorganisms have been and continue to be deeply understudied with many likely having yet to be discovered by modern science since we are still discovering entirely new forms of metabolism and their associated ecological niches and roles which we had no idea even existed all the time in the prokaryotic microcosmos. Some of these have turned out to be major essential roles in the cycling of essential mineral and elemental nutrients particularly in regards to decomposition so we aren't talking about obscure minor players but critical parts of Earth's biosphere of which there absence would have catastrophic consequences as certain waste products would accumulate to dangerous levels.
The two largest bacterial groups which appear to have long specialized in this anaerobic photosynthetic/photoreactive metabolism with facultative and or obligate aerobic metabolisms strategy are the alphaproteobacteria and gammaproteobacteria. Of these notably the alphaproteobacteria are the clade from which mitochondria descend hence they are pretty important all things considered.
Some more potential evidence for this kind of distinction comes from metagenomic sampling which lets us sequence things in an environment without having to culture them in the lab, (which turns out to miss the vast majority of microbes because only a small percentage can actually consume the bovine serum they tend to use to culture microbes), as this has revealed a surprising link between Eukaryotes and particular thus far uncultured Asgard Archaea lineages thus far isolated only from continental environments (soil or freshwater sediments) with aerobic microbes in marine environments turning out to have diverged from continental kin only during the Neoproterozoic (meaning the open oceans were as mineral studies have suggested, likely fully anaerobic up until around ~800 million years ago or so)
In fact given the studies of cyanobacteria Eukaryogenesis was likely restricted to freshwater basins since the cyanobacteria ancestor of chloroplasts is part of a lineage which is obligate freshwater (low salinity) organisms. All together it suggests complex life arose within the continental waters and only far later after over a billion years of adaptation began to colonize the open oceans.
This is where the newest and most familiar tectonic regime comes into play as hydrous mantle plumes turn out to need a sufficiently high saturation of hydrated rocks to accumulate above subducted slabs these magmas which are rich in elements derived from subducted sediments most notably phosphorus thus became able to rise back to the surface in chemically derived mantle plumes. It is at this point that the modern "deep" subduction appears to have begun with the vigorous break up of Rodinia and a profusion of subduction activity as magmas related to supercontinent break up carry the chemical fingerprints of sediment enhanced chemically buoyant mantle.
TDLR the physical evolution of planets may not only be instrumental in shaping planets but we probably owe our existence to it.























")